

12月19日(土) 白山
システム的抑制――人間再生の壁
河本英夫
機能性の出現とともに、機能はそれじたいの自己維持のために、さまざまな仕組みを活用する。肉食動物は、肉食という機能性のために強い顎、短い腸、強くて丈夫な足等々の各器官上の特徴を統合するように働く。機能性(働き)の維持は、生命体の本性であり、いっさいの手段を活用して、その維持を行っているように見える。ところが機能のもとに配置された器官や部位に損傷が生じた場合、機能性そのものは活用できる仕組みのいっさいを活用して、自己維持を図っているようにみえる。そのことが損傷時に多大な負荷をもたらす。
生命の最大で不可欠の機能は、「食べること」と「産むこと」であり、原核細胞のようにただ図体が大きくなってくびれて二つに分かれる場合も、産むことの原初的なモードである。認知機能の基本的な働きのモードを、ピアジェは「同化」と「調整」だと考えてきた。そこから「眼は光を食べている」という名言が生じる。機能性の出現立に、どの程度の働きが関与しているのかは、いまだよく分からない。
抑制的制御は、各局面での機能性そのものの成立に不可分に関与する。そのためほとんどの抑制の仕組みは、生にとってあるいは個体であることにとって「積極的で有効な働き」を行っている。そこには何重にも編み込まれた複雑な制御の仕組みが関与していると考えられる。この積極的な働きが、損傷や解体からの回復の場面においては、突破しにくい壁となる。そこで抑制によるシステムそのものの機構を考察しながら、どのような回路が可能となるかを検討する。
生命の自己性そのものの成立は、一般的にはオートポイエーシスの仕組みによって成立している。この自己性( Selbstheit)に、さらにさまざまな機能性が物の性質として出現する。認知機能も運動機能も免疫機能(自己防御機能)もそうした機能系である。機能は、ひとたび成立すれば、それじたいが維持される。また外的に見ても明白な指標となり、感覚知覚によって観察者も判別することができる。これは質料-形相で見たとき、形相に相当する。形相は、認知の最大のメルクマールである。ところが形相そのものの出現は、いまだ誰も解いていない難題中の難題である。そしてその部分に立ち入らないと、うまく見えてこない事象の領域がある。
一例として、鳥の「飛ぶ機能」を取り上げる。飛ぶための羽は、当初は鳥の体温調整機能として出現し、その器官の可動域を広げて活用していくうちに、たまたま飛べるようになったというのが実情であろう。これは機能転換によって新たな機能が出現した事例である。ひとたび機能転換が起きれば、飛ぶ機能に適合的に起動部位は再編される。この起動部位の一つに損傷が生じた場合、機能維持のためにいままで活用していなかった部位が代替的に組み込まれたり、残存している部位を過活用したりして、機能維持がおのずと図られる。
機能の出現は、起動部位集合から見たとき、比喩的にはハイパーサイクルの出現に最も近い。各部位は、ただ動いているだけなのに、サイクル総体が起動すれば、おのずと機能の維持に参加している。各部分サイクルはそれ固有の作動を行っているだけであり、破損すれば停止するが、にもかかわらずハイパーサイクルはそれを飛ばして作動を維持することができる。また各サイクルの集合そのものを変えることもできる。一部の部位に損傷が生じた場合、それを飛ばしてハイパーサイクルを維持する場合には、滑らかさに欠けてどこか無理が来ていることがあり、代替部位を参加させて、変形した形で機能が維持される場合には、余分な動きが入り込んでいるように感じられる。あるいは機能そのものが停止する場合(歩行の停止その他)が、おのずと出現する。
人間の場合、自分自身の観察者でもあるので、ハイパーサイクルの全体像しか掴むことができない。うまく動けているかどうかという二分法は、観察者の視点から出ている。当人も治療者も、マクロ指標を手掛かりにしている。ところがハイパーサイクルは、それじたいでの動きの判別に対応する構造的要素がない。
ひとたび機能が出現した場合、機能系では、コスト引き下げが決定的である。不要な働きを抑制し、必要な時だけ起動できなければならない。そこには「選択的起動」という仕組みがあるに違いない。たとえば免疫が、自分自身の身体の一部を外敵だと認定して攻撃を開始すれば「自己免疫疾患」となる。身体が必要に対応せず、一定頻度で動きが出てしまえば「多動性障害」となる。各機能は、起動することもできれば起動しないこともあるという選択性をもたなければならない。社会学者のルーマンは、こうした事態を「複雑性の縮減」だと呼んだ。機能系の維持は、おのずとコスト削減の仕組みを備えている。
また一般的には生命とは、放置すれば一挙に進んでしまう反応を可能な限り遅らせる「遅延の機構」の一式である。反応できる作動のモードを抑止しながら制御し、必要に応じてそれを解除して活用する場面に見ることができる。これもコスト削減のモードである。そのとき体細胞系戦略、神経系戦略、免疫的戦略、遺伝的戦略は、それぞれ異なっていると考えられる。体細胞系には、循環器系、呼吸器系、消化器系という主要三機能系が分化してくる。
ここには大別して、(1)総体として未分化なものが発生的、発達的に分化していくが、その途次不要なものを削減していく仕組みがある、機能性とは「捨てる物を捨てて残ったもの」のことである。(2)機能性の出現後に、コスト削減から見て、当面不要な活動を抑止して、潜在性に留め、必要に応じて起動する、これによって潜在性と現動性の区別が出現する。(3)起動可能性に何段階にも作動の度合いを調整できるような段階性が出現する。これは段階的な抑止である。(4)段階区分のための指標が、単独で出現してくる。代表的なものが、快/不快や痛みである。この延長上に「情動や感情」が出現する。情動や感情は、妨げられることではじめて出現する。つまり妨げを感じ取る仕組みであり、妨げられるものと感情や情動の動きには、少なくても一対一対応はない。
1 機能的自己の出現(選択性と調整の出現)
選択性は、生命の基本的な働きの一つである。恒常化された選択性を、人間は「生体の機能」として捉えている。選択性の起源は古く、高分子から出現している。ここに「感覚」の起源がある。カラムにある化学分子を張り付けて、そこに溶液を流すと溶液内の特定の分子だけが結合し、他の分子は流れ出てしまうことがある。分子の特異的結合であり、高分子に出現する特異的反応であり、最初期の認知機能の出現である。溶液が流れ出た後に、カラムに蒸留水を流し、特異的に結合した分子だけを取り出すことができる。この場合には特異的弁別(最初の認知)は出現しているが、いまだ選択性は出現していない。
次に化学反応で、反応産物が一定量になると、反応そのものの速度を抑制することがある。この場合には反応産物は、自分を作り出した反応の速度を調整するのだから、部分的には触媒の働きを行っている。触媒にはすでに速度について「調整」能力がある。一定量の産物ができれば、反応そのものを停止させたり、それが不足すれば再度反応を開始させたりすれば、そこに選択性の必要条件の一つが出現している。反応の起動/不起動の区別が出現すると最初の選択性の出現となる。血糖値が下がれば、シグナルが働き肝臓のグリコーゲンを分解して、血糖値の不足軍を補う。不足を補い、過剰であれば停止する仕組みは、総体としてみれば「動的平衡系」となる。
自己触媒系 「自己触媒システム」の意義は、以下のことである。
第一に自己触媒システムでは、みずからを変数とする関数となるために、ひとたび生成プロセスが生じれば、指数関数的に生成プロセスは進行(遅延)する。
第二に自己触媒システムでは、同一の生成産物が生成プロセスを触媒している。この事態を触媒物質を中心にして考えてみれば、触媒はみずからと同一の物質の産出を指令しているようにみえる。たとえばポリアデニル酸(ポリA)は、ひとたびそれが形成されれば、自己の複製反応をみずから指令すると言われる。このとき触媒は、同一性を支える「情報」をもち、この情報によって生成プロセスが指令されると言われる。「情報」とは、同型の生成プロセスの反復を、静止した「物質」に写像したものに他ならない。
第三にこうして情報担体による触媒サイクルは自働的に生成プロセスの産物を指令する以上、このサイクルの境界は、内的な情報により定められているようにみえる。この段階でコード形成が観察される。コードとは、自己触媒サイクルの生成プロセスの「同一性」を、実定的な規則として静止させたものである。この事態を観察者からみれば、自己触媒サイクルはただコードにしたがって作動しているようにみえる。
この段階が、システムの調整機能の出現であり、この調整機能が情報もしくはコードにしたがって起きているようにみえる。ここでの調整変数は、量と速度である。
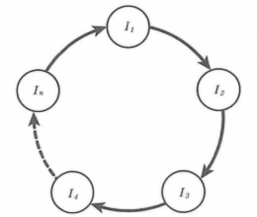
ハイパーサイクル 自己触媒ループのより複雑な機構は、生成プロセスの産物が、次の生成プロセスを触媒し、次の生成プロセスの産物が、次の次の生成プロセスを触媒するように連鎖していき、一つの大きなループをなす場合である。これはより高次の組織化であり、新たな階層段階である。ある生成プロセスの生成産物が次の生成プロセスの触媒となり、こうして順次生成プロセスを触媒することによって、ある段階の生成プロセスの産物が、最初の生成プロセスの触媒になったとき、そこに複合的なサイクルが形成される。異なる機能系の連動が、ここでイメージされていることである。(図1)
複数個の機能系で連動している場合に、一方の系が他方の系の速度調整を行う場合には、連同調性系が出現する。DNAとタンパク系はこうした系で、媒体の主要な作動物質はRNAである。一定量のタンパク質が形成されれば、DNAそのものの作動を停止させる。不足して来れば、再度タンパク質の形成を誘発する。ここにもすでに調整系が出現している。
免疫系 免疫系はそれじたいのなかに弁別反応を備えた認知系である。抗体の蓄積量に応じて、スギ花粉の一定量(抗原)に対して、抗原-抗体反応が起動する。スギ花粉は本来生体にとって外敵ではないので、できれば反応しない方が良い。ところがスギ花粉が免疫システムによって「非自己」の認定を受けると、自動的に起動する。この認定そのものには閾値があり、少数の抗原に対しては起動せず、ある一定量以上の抗原に対しては、起動してしまうような抗体の蓄積レベルがある。
蓄積されている抗体量によって、この起動/不起動の閾値は変動する。ひとたび起動した反応は、放置すれば持続的に作動する。これは免疫システムの行う過剰防衛であり、無駄なコストがかかっている。この段階では、抗原-抗体反応にシステムの履歴と記憶(抗体の蓄積が効いてくるが、ここでは反応することもあれば、反応しないこともあるというような選択性が部分的に出現している。
イエルネのネットワーク・モデルを活用する限り、免疫システムの形成には、ある抗体の出現が、その場所での抗原となり、それによって次の抗体の出現を促し、このパターンが続くことになる。このプロセスは、新たに作られた抗体が抗原になったとき、その抗原に対応する抗体がすでに作られていたネットワーク内に存在する場合には、そこで新たな抗原の出現には当たらない。そのため、ネットワークの制作はひとまず終わりになる。それによって抗体のネットワークは「円環的ネットワーク」となる。人間の場合、約1年ほどでこうしたネットワークが一揃いできると考えられる。
免疫システムへのすべての刺激は、抗原として対処されるのではなく、大半の刺激に対しては抗原としては認知しない。抗体のネットワークは、必要なときには起動し、不必要なもの(害のないもの)に対しては可能な限り起動しないという選択性が当初より必要となる。これは種としての進化をつうじて形成されてきたと考えておいたほうがよい。コスト負担から見て、最も必要とされるのは「免疫的無視」であり、広範囲の刺激に対しては「非自己」認定を行わないことが必要である。免疫的無視は、余分な反応を行わないことである。免疫の認知機能は、「非自己の認定」と「免疫的無視」から成る。
異物や非自己の認定を行うものの集合が、免疫システムの自己すなわち免疫反応を行う一連の細胞のネットワークである。免疫のネットワークによって、異物だと認定されないもののなかには、個体のさまざまなたんぱく質や個体にともにあるような多くの物質がある。口腔には地球の人口ほどの約六〇億個の細菌がある。腸のなかにも膨大な細菌が生息している。そのなかには消化を助ける乳酸菌のようなものもあり、大腸菌も生息している。これらは通常免疫システムによって非自己だとは認定されない。生命体は、多くの体内細菌とカップリングしている。これらは生命体みずからで産出しているわけではないが、連動することで生命機能を維持している。非自己の補集合が、自己になるわけではない。
免疫システムは生体を防御している。だがそれは生体の自己を認知し、それ以外のものを非自己だと認定することによって防御しているのではない。むしろ非自己を認定し、非自己を除去あるいは解除することによって、そのことの副産物のように生体を防御している。そのとき生体そのものにはできるだけ害が及ばないような仕組みを備えているだけである。免疫システムは、栄養源として生体を媒体としている。この媒体のなかではじめて免疫システムは作動できる。
免疫システムは、この点で奇妙な性格を備えている。免疫システムの意味を知ろうとすれば、このシステムが結果として防御している生体そのものと関連付ける以外にはない。だが生体の自己を知り、それを防御しているのではない。免疫システムには、自己を認定する仕組みは備わっていない。システムの作動の結果として生体は免疫システムによって防御される。だが免疫システムの作動そのものは、精確には生体を防御しようとはしていない。そのことの直観的な意味は、生体は免疫システムの媒体ではあるが、いつでも非自己と認定される可能性がある。これが自己免疫疾患で、免疫は自分の媒体を攻撃する。免疫システムに必要なことは、システムそのものが生き延びるために、不用意にみずからの媒体である生体を傷つけないこと、および非自己と認定したものと戦うことであり、この戦いの開始に媒体である生体のシグナルを活用することである。
おそらく免疫システムの進化的な由来は、細菌や異物を食べることであり、分解して栄養の一部として活用することであったと思われる。そのことは細菌をまるごと食べてしまうマクロファージや貪食細胞にそのタイプの働きが維持されていることから推測できる。それが生体にとって、結果として間接的防御にもなる点で、非自己の認定と除去を行うような生体内の機能的組織として複雑化したものと考えられる。非自己の認定と非自己への対応可能性をもつものの集合が、免疫システムの自己である。
免疫細胞の形成 免疫のネットワークの予備集合には、骨髄で形成される免疫細胞の予備軍が含まれる。それは細胞の産出プロセスで供給されるものである。免疫の自己の予備集合もレパートリーを変化させる。この変化のさせ方は、ほとんど環境要因とは独立であり、自分自身の作動のモードにしたがって、おのずと変化させている。
免疫システムは、非自己/非自己でないものの区別のように、二分法の区別を行いながら作動してはいない。法/不法(法システム)、価値/非価値(経済システム)のような区分は、言語の肯定/否定のように世界を二つに分けることを基本的な機能としている。
自己の予備集合は、異物や非自己に対応して形成されるのではなく、ただひたすら形成される。この産出的作動は、オートポイエーシスの一般的な定式化にしたがう。ともかくも産出的作動の本性にしたがって大量に産出される。そして作動機会がなければみずから消滅する。作動機会をもつものは、それによって「記憶」され、潜在的な可動状態を形成する。そうして形成された自己の予備集合は、異物に対して、力学的、生化学的というより、確率的に対応している。膨大なレパートリーを用意しながら、たまたま異物に対応するものが見つかれば、そこで自己の要素が形成され、やがて自己の集合が形成される。そうなると免疫的自己の産出ネットワークは、異物や非自己とは独立に作動し、いわば待ち受けのための網を張っているような状態である。ただしこの待ち受けのための網になるために、何段階かの選択を経る。免疫細胞の分化過程は、免疫的戦士になるための厳しい淘汰段階を潜るようなものである。
免疫的制御 免疫の働きを担う主要な細胞は、T細胞とB細胞である。T細胞は細胞内に侵入するような細菌に対して働きかけ、B細胞は細胞表面に接近してくる細菌や物に働きかける。大幅に簡略化して描いてみる。とりわけ通常は抑制されている免疫細胞が、どのような条件下で起動するかに関心を注いでみる。T細胞の前身の細胞は骨髄中の造血幹細胞から分化し、胸腺の皮質に集められる。胸腺は、実地訓練現場のようなところで、無反応なT細胞やのべつまくなしに反応するようなT細胞は除去される。だから胸腺は、中枢制御室である。胸腺皮質の上皮細胞に発現している生体の主要組織適合性複合体(MHC)分子と弱く反応するT細胞だけが集められ、それらは胸腺髄質において生体由来の抗原の提示を受け、過度に反応するT細胞は取り除かれる。ここで形成される状態が、中枢性免疫寛容である。
この訓練現場で、いくつかの段階を経て、さらに詳細に分化可能な細胞(ダブルポジティブ細胞)となる。この段階でT細胞抗原レセプターが発現してくると、T細胞となることができる。ダブルポジティヴ細胞の寿命は、三,四日しかなく、その間にT細胞抗原レセプターからのシグナルを受け取らないと、ただちに死んでしまう。いわばここではT細胞になるための訓練をしており、この学習を通過しないと、T細胞になることができない。さらに生体由来の抗原やペプチドに対して、弱い親和性をもつものは、デフォルト死を免れ、成熟細胞に分化する。この段階で、生体そのものの固有性との弱い連動を確保し、なおそれに強く反応し過ぎないことが、選択条件になっている。この局面で、生き残るT細胞前駆体は、キラーT細胞とヘルパーT細胞に分化する。
T細胞は、MHC分子に結合したかたちで抗原を識別し、非自己のペプチドを認識して活性化され、非自己抗原を排除するために免疫応答を示す。MHCの遺伝子には、多くの多型がある。MHC分子は本来不安定で、作られてもただちに分解される。そのため生体由来のペプチドを結合したかたちで、細胞表面に発現する。抗体やT細胞抗原リセプターは、遺伝子を繰り返し再構成することで、きわめて多様な抗原に対応する。他方MHC遺伝子は、遺伝子重複と遺伝的多型を獲得することにより、抗原の多様性に対応する。MHC分子は、発現するたんぱく質のなかで際立った多型性があり、これが不一致であると臓器移植のさいに拒否反応が起きる。
胸腺内での選別は、反応可能な範囲の制限を行っているだけであり、何に反応するかを決めているわけではない。しかも選択されれば、それだけで異物と有効に反応できるようになるわけでもない。実際、過敏反応性のT細胞のすべてが胸腺で除去できるわけではなく、五%程度は末端に移動してしまうと言われている。末端に移動した過敏T細胞は、別の機構で起動しないよう制御されている。ここが末端制御機構である。T細胞が作動するために、二種類のシグナルが必要である。これらを欠けば通常激しく反応する抗原に対しても無視してしまう。この二種類のシグナルは、抗原受容体が抗原と結合することで発生するシグナルと、補助受容体やサイトカインからの生理活性シグナルである。二つのシグナルのうちどちらか一方が欠けてもT細胞は作動しない。もうひとつの制御機構は、T細胞の一種である免疫抑制細胞の働きによる。ここには抗原を認定して、抗原特異的に抑制を行うものと、自己反応性のT細胞のアンテナ部分が認定する抑制がある。
他方B細胞も、造血幹細胞から派生、分化してくる。B細胞のもっとも重要な仕事は、外来異物に反応して特異的抗体を産出することである。B細胞表面には、B細胞レセプターが発現してくるが、本体は膜と結合した免疫グロブリンであり、これが外来性抗原を認識する抗原レセプターとして機能する。B細胞も生体由来の抗原やペプチドに反応するようであれば、アポトーシスによって除去されると考えられていた。ところがB細胞リセプターからのシグナルによってB細胞表面の構造が組み替えられことがわかっている。つまり非適合であれば死滅するのではなく、非適合のシグナルがあれば細胞表面のリセプターの構造が組み替えられる。おそらくここにはB細胞リセプターの多様性を確保し、それを拡大していくメカニズムと、膨大な無駄を減らしていく仕組みがあると考えられる。
しかもB細胞の活性化は、通常は抗原との反応だけでは不十分で、T細胞由来のシグナルなどの共刺激抗原とともに働いた場合に、効率よく活性化する。活性化したB細胞は、末梢組織で分化し、抗体産生細胞となる。B細胞は抗原に特異的に反応して増殖・分化し、抗体を産出する。個々のB細胞は、単一の免疫グロブリンを産出するだけであるが、細胞表面を組み替えて、際限のない多様さを形成することができる。これが一般に、免疫システムの多様さと特異さの仕組みだと呼ばれるものである。
抗原レセプターを介するB細胞と抗原との反応は、実は抗原とB細胞との反応だけでは、B細胞の効率的な活性化は起こらない。むしろ抗原との反応だけでは、B細胞はアポトーシスを起こしたり、機能的に不活性化され、免疫寛容が導かれる。いわばロックのかかった状態になる。B細胞抗原リセプターからの反応だけではなく、他の分子からのリセプターを介したシグナルが共存することが必要である。活性化T細胞と同じ抗原に反応するB細胞の接触により、B細胞の活性化が起きる。場所としては、末梢リンパ臓器におもにT細胞が分布するT細胞領域と、おもにB細胞が分布する濾胞領域がある。濾胞の抗原未感作B細胞が抗原に反応するとT細胞領域に移動し、同じ抗原に反応するT細胞と抗原とを介して、活性化する。このあたりは、何段階かにヘッジがかかった仕組みがあることがわかる。こうしたヘッジでは、非自己の認定が直接認知ではないこと、さらにはロックを解除するようにして活性化すること、さらにはT細胞由来の生体そのものの間接的効果も関与していると考えられる。
この段階では部分的に、抗原認定が行われても、動くこともあれば動かないこともあるという仕組みが、成り立っていることがわかる。ここに免疫的選択反応が成立している。認定に複雑な仕組みが関与していることによって、認知系と起動系が分離してきている。認定が行われても、動くこともあれば動かないこともある、あるいは認定が行われてもさまざまな対応がありうるという局面で、認知-実効起動系の選択的制御が出現する。こうした選択性の出現は、観察者から見たとき、「シグナル系」「認知系」と「実効作動系」の分離となり、認知系そのものの出現となる。この段階では認定から起動へとつながる回路に、「選択的抑制」や「選択的興奮」が働いている。この場合認知系と起動系(運動系)は、疑似独立に作動することになり、両者の関係で言えば、「カップリング」である。認知系と実効起動系の独立は、複数の機能系が明確に分離すること、もしくは要素を異にするも複数の機能系が後に連動することで成立する。
感覚の形成 感覚の形成、あるいは五感の形成は、人類史の最大の成果である。発達障害系のうち、アスペルガー、注意障害等は、感覚の抑止が不全であり、あまりにも多くのものに反射反応してしまうモードである。感覚は、無作為に反射反応していたものが、選択的な反応を行うようになることで、システム的な機能をもつ。
感覚が不用意に、また無防備に反応してしまい、余分な反応が起き、身体運動が伴ってしまえば、本人にとってはパニック性の反応となる。その場合、感覚は恒常的に起動可能状態にあるために、身体運動をロックするような場所が快適となる。ソファーとソファーの間であるとか、牛舎での牛の首輪のような外から身体を縛る状態が快適な場所である。(グランディンの事例)感覚が細かく成るという事態は、「抑制的な細分化」によって形成される。つまり「あまり反応しない」「それほど反応しない」「ろくに反応しない」「ほとんど反応しない」等々の抑制的区分によって感覚は微妙さや詳細さを獲得すると考えられる。感覚区分固有領域があり、色は細かく区分され続けるが、音の細かさは形成されないように、感覚区分には個体特異性がある。人間の感情のようなところに細かく区分が働かず、牛の悲しさの度合いには細かく区分が形成される等、感覚の区分には固有領域がある。
強度 そのなかでも世界内の変化率については、なお特異な事態が生じる。背後から猛然と接近してくるトラックの音が聞こえたとき、それに対応する身体反応は、変化率への対応であるが、この対応には一対一対応がない。世界のなかで「変化率」は特殊な事態である。このことをフランスの哲学者ジル・ドゥルーズは「強度」だと呼んだ。
概念はアリストテレスの論理学では、二つの内容をもつ。それが内包と外延である。たとえば「人間」という概念の内包は「理性的であること」であり、外延は人間の集合である。このとき概念からは、個々の人の固有性に届かないことがわかる。中世の哲学者ドゥウント=スコトゥスは、外延は測定可能なもの(メジャーメント)だとし、内包を測定できないが度合いの判別できるもの(強度)に転換していく。その場合、科学とは強度をメジャーメントへの転換する作業となる。お湯の暖かさの度合いは、誰であれ触れればわかる。熱せられた鉄の度合いは、赤、黄、青等々でわかる。そこに基準点を設定して、相対的量の違いで語ることができるようにすることが、科学であり、このとき度合いはある変数の「値」となる。人間の科学的知識は、そのように形成されている。
そのとき度合いの感じ取りと身体運動との関連で、量としてかりに認定したとしても、なお量の認識がそれとして認識に留まらない領域を形成する。たとえば変化率は、変化の変化として量として換算できる。ところが変化率の感知は、身体運動を特定の範囲にとどめることはなく、そこには一対一の対応はなく、また変化率が何であるかを知ることなく身体運動は起きる。変化率は、容易に学習によって制御できない。こうした領域をドゥルーズは強度と呼び、新たなテーマ領域とした。強度は本性上抑制にかからない感覚経験の領域である。
たとえば患者に現れる緊張は、強度の代表的なもので、緊張の度合いは見てただちにわかるが、それを量化することは難しい。にもかかわらず緊張の度合いは、動作ならびに動作の自然性にとって内的に重要である。緊張をゼロor1にしてしまうのは、緊張そのものの本性であり、呼吸法、筋緊張の解除その他をつうじて、会得していくしかない。
強度の記憶は独特である。強度の経験は、異物の経験と同様にコンテキストによる調整を受けにくい。特段の恐怖感であったり、衝撃の経験に類似しており、配置のできない記憶となるが、それ単独で維持されている。コンテキスト化される記憶は制御されやすいが、単独の記憶は、ある意味で「記憶に落ちていない」。記憶‐想起の回路とは別の形で維持されている記憶がある。
触覚 触覚には感覚とは異なる抑制の仕組みがある。反応を制御するのではなく、刺激を感じとらないように反応を停止させるのである。ここが「無視」であり、無視は無駄なコストをかけないための積極的な自己維持の仕組みであり、最少の認知的反応で身体を制御しようとする内的な機構である。掌で机を撫ぜるとき、机の表面のざらつきは2500‐3500程度区分できるが、通常そうした細かな区分は行なわない。触覚の認知は、身体の動作に必要な指標を取り出すだけで、認知的判別が目指されているのではない。それは触覚に身体運動が内在しているからで、認知と身体運動はどのようにしても独立にはならない。
(認知と身体運動が二次的に外的につながる仕組みを考案したのが、ギブソンの生態物理学である。このとき認知的情報は、運動の調整要因として機能する。また身体をともなう知覚についてメルロ=ポンティの身体知覚の両義性という考えがある。手でもう一方の手を触ったとき、能動と受動が入れ替わるという説である。これは身体とともにある知覚が知覚一般として成立することを大前提としているが、手でもう一方の手を握るさいには、前方への運動が含まれており、この運動感はメルロ=ポンティが考えるような反転はしない。また運動感であれば、受動、被動、自動、能動のような細かな区分が成立し、能動-受動のような対関係にはならない。)
ことに身体内感は、身体動作の制御にとって欠くことができないが、個人差も大きく、制御しにくい。身体内感では、麻痺によって位置感覚(ここという感覚)、度合い(力の籠め具合)ともほぼ欠落する。
神経系の形成 神経細胞(シナプス)の抑制は比較的良く調べられている。神経系は、当初ニューロンがともかくも形成され、そこから機能化することによって、機能化しないニューロンが減少することで形成される。ニューロンの総数では、母体内の五か月目ぐらいが最大量で、それ以降は減っていく。それに対して、ニューロン間の接続を形成するシナプスは、胎生8週目ぐらいから誕生後2歳程度まで増えて、さらに同時に胎生16週目から10歳ぐらいにかけて、断続的に神経回路網の再編成が起きる。
この神経回路網の再編には、明確なモードがある。
(1)神経経路網は、当初複数の機能的回路をもつ。たとえば手を動かすときに、その機能に対応する回路は複数ある。これは機能的特定化がいまだ起きていないことを意味する。たとえば1歳程度では、ジャンケンのチョキは周囲の指が一緒に動いてしまい、チョキのかたちがうまくできないことが多い。機能未分化から機能的特定化の方向へ、神経網の再編が生じている。未熟期では、神経系と筋細胞は、複数の神経が一つの筋細胞と接続している。ところが神経系の再編をつうじてその回路網が特定される。このとき複数の回路のうち、特定の回路だけが顕在化し、他の回路は潜在化される、という可能性は、ほとんど成立していない。大脳前頭葉の一部では成立するかもしれない。
(2)神経回路網の再編では、臨界期があり、それぞれの機能に対して、臨界期の時期を過ぎると、もはや再編が起きない。たとえば言語は、生後12年ぐらいが臨界期であり、その時期を過ぎると言語の習得(発語の修得)は行われない。
(3)この再編には、二つの仕組みが関与している。一つは回路網の配線が変わるというものであり、余剰となっている神経回路網が除去されるというものである。この抑制的回路網の除去の分子メカニズムはいまだよく分かっていない。神経細胞のアポトーシスが関与している可能性はある。もう一つは情報の受け渡しのモードがかわるもので、興奮性グルタミン酸とGABA(ガンマアミノ酪酸)があり、GABAは幼少期には興奮性であるが、再編期には抑制性として働くことが知られている。
発達の課題 発達が一つの自己組織化である限り、発達には未決定変数が含まれることになる。発達の機構として実行可能なのは、発達の条件を可能な限り提示することだけである。能力の展開が見られそうな場面で、それがまさに一時的に能力の展開を支えているにもかかわらず、それが次の展開可能性を封じてしまうような局面はいくつもあるに違いない。単純に考えても、たとえば視覚の過度の発達は、触覚性の働きを抑制するであろうし、論理的言語能力の過度の発達は、感情のきめ細かさの形成を抑制するであろう。こうした事態が含まれているために、システムの発達にとっての積極的条件が、どのような課題を生み出してしまうのかは、細かな見極めが必要となる。
機能性の分化のさいに、神経系じたいの生存適応戦略がある。生存戦略はニューロンが他のニューロンと接続することによって作動を継続することだけで規定されている。機能的な神経ネットワークの形成は、新たな回路が確定していくことであり、ここで起きることは「切り絵型」形成である。端子と端子が接続して回路網を形成する場合とは異なり、主要な回路が決まるとその周辺には回路予備網と呼ぶべき未決定なニューロンネットワークが広く残る可能性がある。この部分が、ニューロンの損傷が起きたとき、神経可塑性の可能性を保証していると考えられる。ただし脳内の出血、梗塞のような場合には、この潜在的ネットワークを含めて、機能停止してしまうと予想される。
機能性のネットワークはおそらく恒常的に複数個形成されるが、認知系の場合、作動の速度差によって、主要なもの、付帯的なもののような区分が生じる。また運動系の場合には、実際の出力の選択肢の多さによって、主要なもの、付帯的なものが決まる。つまり認知の作動回路は、認知コストの削減の方向に主要、副次の区分が進行し、動作運動の作動回路は、多様度の増大の方向に区分が進む。ただしこれは生後10歳ぐらいまでで、それ以降はコスト削減を基調とする。
新たな機能性の獲得は、一般に関数の比喩で語れば、新たな変数の出現する事態に喩えることができる。それは神経ネットワークでは新たな回路が形成されることと同じだが、新たな変数の獲得は、既存のネットワークの再編を含むと考えられる。この再編は、独立機能領域を新たに領域化する場合には、既存の機能領域とのカップリング関係を再編し、また既存領域の高次化の場面では、既存の機能領域の再組織化を含む。1歳から7歳までの歩様の再編は、事実である。この場合、抑制は、ほっておけばおのずと進行する事態に遅れと選択の幅を作ることである。
そこに多くの代償機構が出現する。(a)本来並行的である二つのシステムのうち、一方の作動速度もしくは作動の駆動要素が一方のシステムによって規定され、他方はただ従属するだけになる。この場合には、一方のシステムの作動可能性の範囲に、他方のシステムが配置されるだけになり、過剰展開と過小展開が現実の形となる。たとえば言語系のシステムと感覚のシステムで、言語からしか経験が展開しないのであれば、言語に対応する感覚的な経験しか分節しなくなる。(システムの従属) (b)一つのシステムが現実の多くを制御できるようになると、他のカップリングしているシステムを実質的に解除してしまう。つまりなんらかの連動関係は残るものの、他のシステムの作動・展開を実質的に無関心的なものとしてしまう。あるいはそうした連動モードになるように、特定のシステムだけを機能強化してしまう。サヴァン・イディオットのような多くの特殊能力は、カップリングの実質的無効化をともなう。さらに麻痺の場合には、代償機構をむしろ積極的に活用しながらシステムの展開がみられる。
2 システムの自己維持――損傷に対して
歴史的事実としては、18世紀末から19世紀初めにかけて、「生物学」が誕生したとき、中心となった概念に「有機構成」がある。「体制」「有機体制」その他多くの語で翻訳されているが、原語は「オーガニゼーション」である。この時代の流行語だったと考えてよい。
有機構成にさまざまな内容を盛り込み、そこで取り出されたものの多くは、動的自己維持の仕組みである。「動的平衡系」がさまざまなかたちで取り出されている。この動的平衡で維持されている当のものが「機能」である。動的平衡系は、ダイナミックな抑制系(補填系)でもある。しかもこれは「構造的な抑制の仕組み」としても機能する。
*生物で「種」の変異(偏移)が問題となるが、容易に次々と変異していくなら、種という固定がない。(ラマルクの場合)種の変化という事態が成立する場合には、種の安定性がどこかで確保されていなければならない。この動的安定性をささえるものが「有機構成」である。
体細胞的損傷への対応 1)損傷したとき、周辺の器官が機能の代行を行い、機能代替が出現する。胃潰瘍で胃の大部分を切り取った場合、12指腸の末端や食道の末端が消化・吸収の代行を行うようになる。器官の相互位置は不変で、欠落は隣接する部位の変化で対応する。(相互補填の原理)
2)特定の部位が膨張したとき、隣接する部位の縮小が起きる。(平衡の原理)これによって同じ有機構成をもつものが、多様な形態をとることができる。
3)各器官の協同の原理――肉食動物の場合、一連の特徴的な器官が生じる。顎が強く、腸が短く、足がど太くて短い。特定の目的に適うように一貫した再編を受けるために、器官の間では目的の維持に沿って、損傷部位の各種補償が起きる。
4)筋膜のような組織は、器官によって異なった外見をもつが、炎症時には類似した変化が生じる。これらは膨張増大・縮小減退を前提とし、有機構成の維持、動的平衡性の持続を大前提としている。
5)動作のバランスが崩れているときには、どこかが弱り、その分を代償的に補っていることが普通である。「力学的有機構成」と呼ぶべきものがあり、歩様の変容は外的見てくれの是正ではなく、部分間の自己維持のバランスを変えていくことが必要となる。このバランス変換をどこに介入して実行するかで、セラピストの腕に違いが出る。(ここが名人芸からただの事務職員的セラピストまで別れる場面である。)
損傷後には自己維持の仕組みが自動的に起動しており、それはきわめて強力であり、介入のコンテキストをさまざまな角度からさらに広く取る必要がある。
5-1)機能変換――歩行不全のような機能不全の場合、歩行を回復させるのではなく、歩行の手前のバランスを変容させる。たとえば鳥の羽は体温調整として機能していたが、やがてそれが飛ぶための器官となる。飛べない鳥を飛ぶことに向けてエクササイズを行うのではなく、体温調整機関として活用することの延長上に、副産物として別の機能が出現する方向での治療がありうる。器官の再機能化と呼ぶべきプロセスがあり、機能を維持するのではなく、再度機能そのものを立ち上げるような仕組みを考案する。病前の状態に戻すのではなく、また病前の状態を目標とするのではなく、治るとは治療行為の副産物であるという点に力点を置く。もっとも近い感覚は、歩行の訓練を行うさいに、歩こうとするのではなく、別の訓練をやっている間に、おのずと歩けるようになるという事態をどのようにして成立させるかである。
5-2)起動部位集合のリセット――疾患後に活用していない部位、もしくは回復過程で動作の起動集合に含まれない部位を、意図的に動作に参加させる。「起動部位集合のリセット」と呼ぶべき事態で、現状の集合のあり方を人為的に変更し、その後エクササイズのなかで本人自身が動作訓練をつうじて、再集合化をおのずと実行する。このときあまりにも広い集合だと効果が薄く、狭すぎればリセットにならない。 こうした範囲を、「介入の最近接領域」だと呼んでおく。発達の最近接領域は、周囲の人の手を借りれば実行でき、後に一人でも実行可能になるスキルの領域(ヴィゴツキー)だが、それに合わせて介入の最近接領域を設定できる。疾患前に活用していた動作集合のモデルが、少なくても小脳には残っているはずで、このモデル的設計図と起動部位の集合がずれていることが普通である。その場合、モデル的設計図に合わせようと無理な起動部位集合を活用していると考えられる。
5-3)「カップリングの活用」駆動部位の周辺には、連動している筋群があり、起動部位だけに働きかけると緊張が出る場合には、連動する部位を動かし、それを触媒のように使って、起動部位集合の動きを滑らかにする。
神経系損傷への対応
1)脳神経系の損傷では、その部位近傍からの情報は、半ば不可避的にノイズを含む。そのノイズをブロックするようにして機能抑制が起きる。骨折、関節疾患でも、その部位を動かさなくなることと類似的である。損傷部位のブロックと同様に、その部位の機能を別の部位で補うような機能代替は、同時に急速に進む。だが体細胞、骨格系の損傷とは異なり、自然治癒はないと考えられる。神経系の自発的再生は、ほとんど考えられない。
2)既存の神経系回路のブロックの破損――手を前に出そうとして肘を伸ばそうとしたら、方が前に出てしまうような場合には、動作の起動部位回路の変容が起きてしまう。動作順序の組み立てができないか、一部回路が切断されている可能性がある。
3)損傷部位の機能を隣接する部位を活用して、あるいは脳対側を活用して、対応しようとすることによって、損傷部位の抑え込みを行う。これは通常訓練を行えば、脳対側での代用が進行し、脳のレベルで機能変換を行うことを意味する。(そのため脳対側を非活性化して訓練を行うやり方を採る人もいる。)
4)神経系の再生をめざして機能的訓練を行い、新たな神経回路が形成されるさいには、(1)すでに形成されていて、潜在的な活動に留まっている回路を浮かび上がらせ、現実化する、これは損傷範囲がごく局所的な場合のみ可能である。(2)新たな神経回路の形成にさいして、損傷時は複雑な回路が形成されると予想される。利用可能な隣接回路を探し出すが特定の機能だけが形成されることはない。歩行時に手が動いてしまうとか、頭に力が入るとか、動作バランスの維持に動作にほぼ不要な動作単位が入り込むことが起こる。(3)特定の機能を再度形成しようとすれば、機能特定は残存する神経回路網をさらに減少させることで実行することになると予想される。
認知的動作誘導は何を行っているのか
1)動作に注意を向けることで、起動部位に出やすい反射的緊張を抑制する。
2)通常は意識以前に進行している動作を、意識の管理下に置くことで、一つ一つの動作の区分を行いながら進行させる。(自宅に帰れば、こうした動作は通常は行わなくなる。) 局所的に認知を向けながら、部分動作を形成するさいには、動作総体の組み立てにいたらないために、現実の動作遂行が不調に終わる可能性が多い。局所の動作単位の回復から、全身的な動作は、多くの選択肢がありすぎてそこをつなぐ訓練が必要となる。
3)動作の調整のための変数を多くする。歩行時には、方向を調整するために3,4メートル先の指示点を確保しながら動作を行う。
4)個々の動作の動作内感を呼び出そうとする。
5)かつての動作のイメージを活用しながら、動作のかたちに自分の動作を沿わせるように実行する。外的なかたちに沿うように動作を作り出すことになる。
6)認知的指示を行うことで、神経系の起動部位の範囲を拡張し、そのなかで動作に対応する神経系の再集合化を行う。この場合最終的には、認知が介在しなくても自動的に動作が進行するようにさらに再編が必要となる。
意識は促進的か抑制的か
意識の機能 意識は、情報処理が間に合うほどの保持時間をもたらすものであり、おおざっぱな認定を行うようになる機能だとされている(コッホ『意識の探究』)。
意識の機能性:意識の機能性を書き出すだけであれば、かなりたくさんのことを取り出すことができる。短期記憶へのアクセスの促進、知覚したものの分類、意志決定、行動の計画、動機づけ、複雑な課題の学習、問題の検出、現在という時の指標づけ、トップダウン型注意、創造性、再帰性モデルの作成、推測、推理のように意識が関与していると思われる働きを取り出すことができる。だがいずれも機能として設定されていれば、意識がそれとして出現する以前に意識下でそれに対応する「ゾンビ・システム」が作動しているはずである。
意識は実に多くの働きをしている。意識は、脳神経科学の見通しで考えれば、大脳前頭葉の発達にともなって、意図せず出現してしまった副産物のようなものであろう。だがたとえ意識が副産物の結果だとしても、自己組織化の本性上、ひとたびある水準の仕組みが創発してしまえば、創発の後にはまったく異なる局面が生じる。意識の出現は、一つの相転移であるが、相転移以前の状態をどのようにしても知ることのできない相転移である。この相転移は、不連続な飛躍を経るだけではなく、飛躍以前の状態が一切解消されるために、みずからの由来を問うことができない。つまり意識は、みずからがどのようにして生成してきたのかをみずからの内部をいくら調べても取り出すことができない。
少なくとも意識には、みずからの活動をそれとして感じ取ることができ(アウェアネス)、みずから自身を知ることができる(自己意識)。それとして意識の自己を感じ取るアウェアネスが、認知科学で「クオリア」と呼ばれたものである。またみずからを一つのまとまりとして感じることができ、みずからの前史を解消し、さまざまな情報に対しての選択的制御、記憶されるものとそうでないものの選択的制御、さらにはみずからの活動への強弱の制御(集中したり、緊張を少し緩めたりという制御)のような働き、そして意識は意識以外のものへと向かうという働き(志向性)を行っている。0.5秒以上持続しない経験は、意識の表象にも意識でそれとして気付かれることもない(リベット『マインド・タイム』)。変化を経験させるとき、0.5秒以上続く変化の開始、変化の持続、変化の停止、それとして経験したものがもっとも記憶に落ちやすい。
意識の組織化機能は可能か:(1)意識が出現することそのものに同時に伴う組織化、あるいは神経システムの組織化の一面が意識の出現であるような事態を想定することはできるか。(2)意識が出現してのちの調整・制御機能の組織化、これはゾンビ・システムそのものにも働きかけているために、意識の働きはどの程度まで及ぶのかが問題となる。(3)意識がそれとして自覚的になることにともなう、ゾンビ・システムの自己維持・抑制、あるいは意識そのものの自己維持によるゾンビ・システムの発達停止がどの程度の範囲に及ぶのか。
ある身体動作ができるようになったとき、あるいは他動的に動作を実行したとき、それが意識される(気づかれる)「速度」を探り当てること、同時にイメージを活用すること、筋緊張の解除と意識された動作にすること、十分に意識が向くようになったら動作の向くさきに注意を向けること(方向づけ)、意識システムがすでにゾンビ・システムを制約している可能性のある場合は、もっとも浮かびやすいイメージ(抜けるような青空、海、地平線等)を設定して動作を行うことも選択肢である。
1)意識的注意を向けて動作訓練を行ったとき、その後同時に別の場所に注意を向けて同じ動作を繰り返すこと
2)意識的注意を向けて動作訓練を行ったさいには、意識の追跡速度を超える速度での同じ動作を行うこと(追跡速度は、年齢、病態によって異なる)。
